Densidad de biomasa aérea en bosques extensos del Neotrópico húmedo. Caso de la Cuenca del Río Caura, Venezuela
Resumen
Por su importancia como reservorio de carbono, se planteó cuantificar las densidad de biomasa aérea en los bosques de la Cuenca del Río Caura (aproximadamente 45.000 km2) al sur de Venezuela. Tomando como base información de estudios realizados en bosques de la Cuenca, se elaboró una base de datos (218 parcelas, superficie 120 ha), con las variables ubicación, clima, tipo de bosque, área basal y volumen en 7 zonas geográficas de la Cuenca.
Como una primera aproximación, se estimaron a partir de datos de volumen de fuste los valores de densidad de biomasa aérea árborea (BA) de los bosques regionales aplicando las ecuaciones alométricas desarrolladas por Brown (1997) para bosques húmedos tropicales densos.
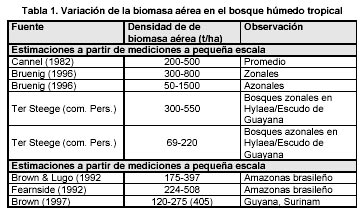
El valor promedio de BA del bioclima húmedo tropical (BHT) en el conjunto de zonas geográficas fue de 237 49 t/ha. La menor biomasa promedio se presenta en las zonas con los suelos más pobres desarrollados a partir de rocas de la Provincia Roraima. El rango obtenido se corresponde con intervalos estimado de BA para bosques zonales del Trópico húmedo de 200-500 t/ha (Cannel 1982); también es similar a la BA de 227 t/ha (166- 332), estimadas con datos del inventario RADAM en el Amazonas brasileño (Brown & Lugo 1992).
Según los tipos de bosques, el promedio de BA en los bosques medios densos (BMD) del bioclima húmedo tropical es de 234 58 t/ha, superior al mostrado por el bosque bajo denso (BBD) de 212 20 t/ha y similar al promedio 239 41 t/ha del bosque ribereño. En cuanto a los extremos inferiores, se puede apreciar que el menor valor de BA del BBD (197 t/ha) y de los menores valores de BA del BMD (205 t/ha) se presentan más frecuentemente en la zona con los suelos de menor fertilidad de la cuenca. El rango de valores de biomasa aérea obtenidos para los distintos tipos de bosques de la cuenca está dentro del amplio rango estimado de 50-1500 t/ha para bosques azonales y zonales del Trópico húmedo (Bruenig 1996) y de 69-550 t/ha para bosques del Escudo de Guayana.
Finalmente, estos resultados de BA obtenidos con el método Brown (1997) se compararon con las estimaciones de BA obtenidas por estudios a pequeña escala, donde se aplicó el método de cosecha destructiva (Dezzeo et al. 1993), constatándose las diferencias entre los resultados de ambos métodos. Por tal motivo, a los resultados obtenidos por el método Brown se le aplicaron las correcciones propuestas por Fearnside (1992) para dicho método. Las estimaciones ajustadas así obtenidas se acercaron a los valores estimados a pequeña escala. Es evidente la necesidad de mejorar los niveles de precisión en la estimación de biomasa aérea en bosques del Caura. Para lograr tal cometido un necesario próximo paso es el desarrollo de ecuaciones alométricas adaptadas a los bosques regionales a partir de información de biomasa colectada directamente en el terreno.
Introducción
La biomasa aérea arbórea se estima usualmente mediante la aplicación de ecuaciones de regresión alométricas a un conjunto de árboles de una parcela medidos, partiendo de la cosecha destructiva de árboles individuales seleccionados o el conjunto de árboles en la parcela. El desarrollo de ecuaciones alométricas para árboles en sitios de bosques específicos requiere de una labor intensiva, por ejemplo la cosecha de un árbol emergente puede a requerir varios días de trabajohombre (Clark et al. en prensa). Por tanto, los investigadores utilizan comúnmente ecuaciones alométricas ya existentes globales o particulares para otros sitios ( Brown 1997). No obstante, es necesario considerar que el potencial de variación entre sitios forestales en aspectos como diversidad de especies, arquitectura arbórea y densidad de madera es grande, por lo cual la práctica anterior puede incurrir en errores (Clark et al. en prensa). De acuerdo a lo anterior, según la intensidad de trabajo y el grado de precisión se podría hacer una distinción de dos enfoques para la estimación de biomasa, el primer enfoque se refiere a aquellas estimaciones realizadas a partir de mediciones a pequeña escala, es decir mediciones directas y costosas, donde se desarrollan ecuaciones alométricas locales, y el segundo enfoque se asocia con las estimaciones a gran escala aplicando ecuaciones alométricas globales.
Es evidente que la estimación directa en el terreno de biomasa de bosques tropicales es una empresa costosa y exigente, tal costo y esfuerzo se multiplica si se trata de áreas extensas, remotas y poco accesibles como es el caso de la Cuenca del río Caura. Esta cuenca hidrográfica comprende una extensa área que abarca aproximadamente 45.336 km2, cubierta en su mayor parte por bosques. El presente estudio tuvo como fin cuantificar a nivel exploratorio y preliminar la densidad de biomasa aérea en bosques bajo los distintos bioclimas presentes en la Cuenca del Río Caura al sur de Venezuela. Tal cuantificación suministra infomación básica que permite evaluar el potencial e importancia de dichos ecosistemas como reservorios de carbono atmosférico.
Densidad de biomasa aérea a pequeña y gran escala en bosques húmedos tropicales
La enorme diversidad y variación de los bosques tropicales y la gran variedad de enfoques y métodos existentes para estimar biomasa hacen difícil una comparación de los estudios en distintas regiones boscosas tropicales. A pesar de dicha dificultad, en la tabla 1 se presenta una síntesis de diferentes estimaciones de biomasa en bosques húmedos tropicales tanto del Trópico húmedo como del Escudo Guayanés y el Amazonas. En la misma se diferencian dos grupos. estimaciones a pequeña escala y estimaciones a gran escala. En dicha síntesis es posible apreciar la enorme variablidad en la biomasa de los bosques; la biomasa aérea total oscila usualmente en el Trópico Húmedo entre 200-500 t/ha (Cannel 1982), pero al incluir los extremos se amplia notablemente el rango para los bosques zonales (300-800 t/ha) y para los azonales (50-1500 t/ha) (Bruenig 1996). Las mayores acumulaciones de biomasa en el Trópico Húmedo se registran en los bosques de Dipterocarpaceae en el Sudeste asiático, mientras que en el Neotrópico son menores (Bruenig 1996). Bosques sobre suelos muy pobres o inundables tienen usualmente baja biomasa (Cannel 1982). En la Hylaea amazónica y la Guayana los mayores valores de biomasa se alcanzan en lugares con condiciones edáficas relativamente buenas, mientras en bosques con suelos extremadamente desfavorables como es el caso de la Caatinga Amazónica la biomasa tiende a ser inferior (Jordan 1989, Bongers et al. 1985, Klinge & Herrera 1983, Klinge & Medina 1979, Klinge et al. 1975). Si se contabiliza el conjunto de bosques del Escudo Guayanés se detectan dos grupos de bosques. los bosques altos zonales con 300-550 t/ha y los marginales con 69- 220 t/ha.
 |
|---|
Area de estudio
La cuenca del río Caura se extiende entre las coordenadas 3° 37’ – 7° 47’ Latitud Norte, y 63° 23’ – 65° 35’ Longitud Oeste, ubicándose al sur del río Orinoco, en el Estado Bolívar.
El río Caura con más de 700 km. de longitud, y un aporte de 3.500 m3/s de agua al río Orinoco, es el segundo afluente más importante del río Orinoco y la tercera cuenca en importancia del país. La cuenca del río Caura registra un gradiente pluviométrico que varía entre 1.300 mm anuales hacia el norte, hasta 4.000 mm anuales en la cuenca media del río Erebato, con una temperatura promedio que varía entre 27°C en las planicies del Sipao al norte y hasta 14°C en la meseta de Jaua al sur (Rosales & Huber 1996).
La fisiográfía es diversa y comprende una planicie aluvial al norte a unos 50 msnm de altitud, una peniplanicie suavemente ondulada y lomeríos en la sección central de tierras bajas entre 50 y 500 msnm, las grandes altiplanicies o “tepuis” al sur entre 1.500 y 2.350 msnm, las montañas bajas en los límites sur y sudoeste de la cuenca entre 500 y 1.000 msnm y los altos macizos montañosos en el límite oeste entre 1.800 y 2.400 msnm. La Cuenca del Caura puede dividirse en 3 secciones. El Bajo Caura; desde la confluencia con el río Orinoco hasta el Salto Pará. el Medio Caura; desde el Salto Pará hasta la confluencia con los ríos Merewari y Waña, y el Alto Caura; desde la confluencia de los ríos Merewari y Waña hasta sus fuentes en las Sierras de Uainama y Aribana (en la frontera con Brasil). El desnivel entre el nacimiento del río y su desembocadura en el río Orinoco es superior a los 2.000 m (Rosales & Huber 1996).
La cuenca se encuentra asentada sobre el escudo precámbrico de Guayana, diferenciándose 4 provincias geológicas, la mayor parte de la Cuenca se asienta sobre la provincia de Cuchivero; en la base nororiental se presenta la Provincia de Imataca y en la parte centro occidental destaca la Provincia de Pastora del Caura; finalmente al sur se presenta la Provinicia de Roraima (Rincón & Estaga 1996). Los suelos del orden Ultisoles son los de mayor extensión en la cuenca, seguido por los Entisoles y en menor extensión los Oxisoles, Inceptisoles e Histosoles. La mayoría de los suelos de la cuenca son fuertemente meteorizados, ácidos y de baja fertilidad (Fuentes & Madero 1996).
En la cuenca se presentan formaciones vegetales de bosques, sabanas, herbazales y arbustales. Aproximadamente el 90 % de la cuenca está cubierta por vegetación boscosa, que en su mayor parte es de carácter siempreverde.
Una estimación conservadora permite afirmar que hasta la fecha menos de un 5% de la superficie total de la cuenca del río Caura ha sido reconocida en estudios científicos. Esto pone de manifiesto la insuficiencia del conocimiento sobre esta región que es considerada como una de las últimas fronteras forestales intactas del mundo tropical (Miranda et al. 1998).
Se han realizado diversos estudios sobre la vegetación boscosa de la cuenca del Caura (Rosales 2000, Knab- Vispo 1998, Castellanos 1997, Briceño et el. 1997, Dezzeo y Briceño 1997, Huber & Rosales 1997, Huber tel. 1997, Rosales et al. 1997, Salas et al. 1997, Marín y Chaviel 1996, Rosales & Huber 1996, Aymard et al. 1997, Briceño 1995, Díaz & Pernía 1995, Dezzeo et al. 1993).
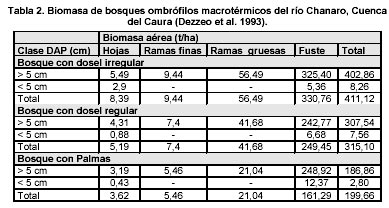
Previo al presente trabajo, en el Caura solamente se disponía de estimaciones de biomasa de bosques ombrófilos macrotérmicos provenientes de un estudio a pequeña escala (Dezzeo et al. 1993) realizado en una única localidad de la cuenca del Caura, el Río Chanaro, el cálculo se hizo a partir de cosechas destructivas en el terreno y el desarrollo de ecuaciones alométricas locales. En dicho estudio se seleccionaron tres tipos de bosque, levantándose una parcela en cada tipo de bosque (ver Tabla 2). Las estimaciones hechas para el bosque con dosel regular (bosque bajo denso) y el bosque con dosel irregular (bosque medio denso) se corresponden con el rango de biomasa medidos a pequeña escala estimado para bosques altos del Escudo Guayanés y Amazonas entre 300-550 (Tabla. 1). Ello no ocurre con el bosque con palmas; sin embargo este último tipo de bosque si se corresponde con el rango de los bosques marginales con 69-220 t/ha (Tabla. 1 y Tabla.2).
 |
|---|
Unidades de vegetación
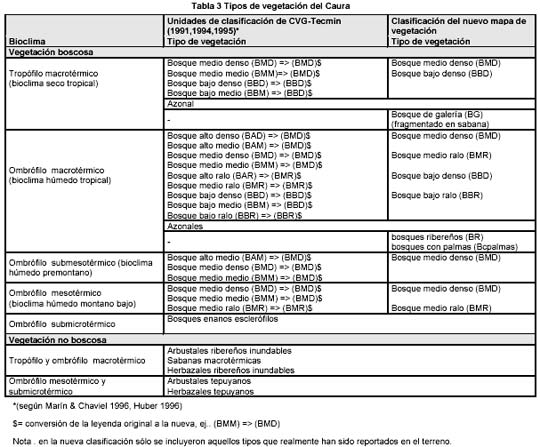
Las unidades de vegetación seleccionadas para el presente trabajo fueron establecidas a partir de una modificación (ver Tabla. 3) de la leyenda de unidades cartográficas tomadas del mapa de vegetación de CVG-Tecmin (1995, 1994, 1991) de la
cuenca del río Caura (ver mapa 1).
Mapa 1. vegetación de la cuenca del Caura (tomado de Marín y Chaviel 1996)
Estimación de biomasa aérea
En una investigación de tipo exploratorio y preliminar como la presente y realizada en un área de estudio extensa y poco accesible, era necesario que la estimación de biomasa no incurriese en costos elevados, tales como los que exigen estudios detallados a pequeña escala mediante cosechas destructivas en el terreno, en donde a causa de la elevada intensidad de trabajo, el tamaño y el número de parcelas o unidades de muestreo a medir serían necesariamente reducidos.
 |
|---|
Una segunda alternativa, consiste en una estimación de biomasa de tipo indirecto a partir de una base de datos dasonómicos a gran escala, alimentada por información preexistente generada por diferentes estudios e inventarios de bosques realizados en la cuenca.
En la primera alternativa, hay que tener en cuenta que los estudios de biomasa a partir de mediciones de un pequeño número de parcelas en el terreno pueden difícilmente lograr cubrir la heterogeneidad típica de los bosques húmedos tropicales, además se presentan limitaciones en cuanto a su aplicación extensiva a otras localidades, más aún si se pretende
extrapolar a grandes superficies. Asimismo, según Brown & Lugo (1992) los datos de biomasa estimados a partir de un reducido número de pequeñas parcelas, restringidas a una o pocas localidades geográficas tienden consistentemente a sobrestimar la biomasa. Por lo tanto, el grado de dependencia de las estimaciones de este tipo de estudios afectará en el mismo nivel nuevas estimaciones que se realicen tomándolas como base (Brown & Lugo 1992). La información sobre cantidades y estructura de la biomasa estimadas en parcelas pequeñas solamente podría ser extrapolada si su posición de muestreo en relación al patrón de variación espacial y temporal del bosque circundante es conocida (Bruenig 1996).
En cuánto a la segunda opción, la estimación de biomasa total mediante mediciones a gran escala en el terreno, la situación tampoco es sencilla. La obtención de estimaciones confiables basadas en mediciones totales en grandes extensiones de bosques exigiría enormes sumas de dinero y de tiempo. Afortunadamente, las relaciones alométricas entre diferentes partes arbóreas se conocen bien y son relativamente constantes y conocidas para áreas vecinas. Es posible por tanto estimar la biomasa de un árbol a partir del conocimiento de variables como diámetro a la altura de pecho (DAP), relación DAP-altura, área basal y volumen de fuste (Brown 1997, Cannel 1982, Hase & Foelster 1982), dichas variables son usualmente medidas en inventarios forestales a gran escala y en estudios botánico – ecológicos a pequeña escala. El análisis de estos argumentos permite concluir que esta última opción es la más apropiada para el logro de los fines del presente trabajo.
En este sentido, tomando como base la recopilación y organización de información proveniente de 22 publicaciones e informes inéditos (ver en bibliografía las referencias marcadas con *) sobre bosques de la Cuenca del río Caura, se elaboró una base de datos a nivel de parcelas con las siguientes variables. clima, ubicación geográfica, paisaje, tipo de bosque, área basal y volumen (cuando éste dato estaba disponible) de las diferentes zonas boscosas de la cuenca, utilizadas posteriormente para la estimación de la densidad de biomasa aérea de los bosques regionales. Tal base de datos se basa en información acopiada en 218 parcelas que suman 120 hectáreas de bosques, esta base ofrece una cobertura espacial geográfica y climática de gran parte de la cuenca, lo cual posibilita la comparación de las diferentes zonas geográficas y tipos de bosque así como una visión de conjunto. La principal fuente de información es la proveniente del inventario de recursos naturales de la Guayana Venezolana (CVG-TECMIN 1991, 1994, 1995). Una de las principales limitaciones tanto del
inventario de CVG-Tecmin como de los estudios restantes usados como fuente de información consiste en que la selección de los sitios de muestreo dependía en alto grado de la accesibilidad y los recursos disponibles; esto naturalmente determina un sesgo en el muestreo.
 |
|---|
La estimación de densidad de biomasa aérea se basó en valores de volumen de fuste. En el caso de que se tratáse de parcelas sin datos de volumen, dicho valor fue estimado a partir de una ecuación de regresión entre área basal y volumen en aquellas parcelas, donde estos datos estuviesen disponibles. Al respecto, es necesario recordar que el área basal arbórea es una medida bien fundamentada y fiable para estimar biomasa de bosques tropicales (Philips et al. 1998).
El método utilizado para la estimación de biomasa es un modelo desarrollado para bosques tropicales (Brown 1997, Brown & Lugo 1992, Brown, Gillespie & Lugo 1989, Brown & Lugo 1984).
Está basado en el uso de datos preexistentes de volumen por hectárea, y se puede aplicar de manera general a bosques secundarios y maduros, presentes desde climas húmedos hasta secos. Sin embargo, lo más apropiado es usarlo en bosques densos, ya que los datos originales usados para desarrollar el modelo provienen de tales tipos de bosques (Brown 1997).
La densidad de biomasa aérea fue estimada mediante el uso de la siguiente fórmula (Brown 1997, Brown & Lugo 1992, Brown, Gillespie & Lugo 1989, Brown & Lugo 1984).
BIO = VOL*DM*FE
BIO=densidad de biomasa aérea, entendida como el total de materia orgánica arbórea viviente, expresada como peso seco al horno en t/ha VOL= Volume de fuste en m3/ha de todos los árboles > 10 cm de DAP (ver anexo 5)
FE= factor de expansión de biomasa (proporción o cociente entre biomasa aérea seca de los árboles y biomasa seca del volumen inventariado), en donde.
FE= 1,74 si BV> 190 t/ha, si BV< 190 t/ha, entonces FE= EXP ((3.213- 0.506*ln (BV)),
BV= biomasa del vol. inventariado en t/ha, calculado como VOL/ha (m3/ha)* DM (t/ m3)
DM=densidad seca de la madera= 0,66 t/ m3 o g/cm3, estimado como valor promedio de la densidad seca de madera de las especies más importantes de bosques en la Cuenca del Caura, los valores de DM por especie fueron tomados de la literatura
(Brown 1997, Fearnside 1997, JUNAC- PADT. REFORT 1981, Gerard et al. 1996). Este valor es similar al estimado (0,67 t/ m3) para bosques densos en la vecina región de Roraima en el amazonas de Brasil (Fearnside 1997). El valor promedio de densidad de la madera para árboles del Neotrópico ha sido estimado en 0,62 ± 0,01 t/ m3 (según Chudnoff 1980, cit. por Brown & Lugo 1984).
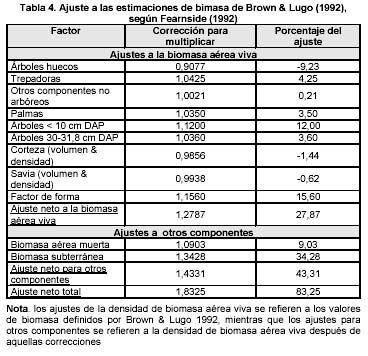
Este modelo de Brown ha sido objetado por Fearnside (1992), quién argumenta que dicho método subestima la biomasa. Por lo tanto, él propone corregir dicha subestimación aplicando unos factores de corrección (ver Tabla. 4). Una vez hechos dichos ajustes, los nuevos valores corregidos de densidad de biomasa aérea se incrementarían acercándose a valores similares obtenidos en estudios de pequeña escala (ver Tabla. 1). Se considera que se puede hacer uso de ambos enfoques teniendo en cuenta las diferentes metodologías de estimación cuando se hagan comparaciones. La ventaja del modelo de Brown es el poder hacer uso de información preexistente, su menor costo y su aplicabilidad a estudios de gran escala como el presente, aspecto que incluso Fearnside (1992) reconoce.
Resultados
En esta sección de resultados se hará énfasis en los bosques ombrófilos macrotérmicos, por ser la formación boscosa con mayor extensión superficial en la cuenca, en consecuencia la que representa el mayor reservorio de carbono.
Los bosques neotropicales son heterogéneos y el conjunto aquí presentado abarca mucho de la variación natural de los bosques del Escudo de Guayana. Los agentes determinantes de tal variación de la biomasa en los bosques tropicales aún no se han dilucidado completamente; sin embargo es posible decir que el tipo de suelo es uno de los principales factores determinantes. Lamentablemente, no se dispone aún de información sobre la relación existente entre tipos de suelos y tipos de bosques para la mayoría de las parcelas que alimentan la base de datos. Por lo cual, el suelo no será considerado como un parámetro en la estratificación y análisis de los bosques, presentados a continuación.
Biomasa aérea, bioclima y zona geográfica
Las zonas geográficas (definidas por MARNR 2000) utilizadas en la estratificación de los bosques se muestran en el mapa 1.
En la tabla 5 se puede apreciar la densidad de biomasa aérea de los bosques estudiados según el bioclima y zona geográfica. En la Tabla 6 se presentan los mismos parámetros pero discriminados según los tipos de bosque.
De la tabla 5 se desprende que el valor promedio de la densidad de biomasa aérea de los bosques en el bioclima seco tropical (BST, sensu Holdridge 1947, equivalente al bosque tropófilo macrotérmico) no se diferencia claramente de la biomasa boscosa del bioclima húmedo tropical (BHT, equivalente al bosque ombrófilo macrotérmico). La causa de dicho fenómeno se explica al analizar la distribución de la biomasa aérea de los diferentes tipos de bosque en el BST (Tabla. 6), donde se observa que los bosques azonales de galería presentan una mayor biomasa que los bosques de interfluvio; muy probablemente tal biomasa mayor es producto de las condiciones edáficas particulares de los bosques de galería en una zona bajo un clima con un período de sequía bien marcado.
Por otra parte se observa que, la biomasa del bioclima muy húmedo montano bajo (BMHMB, equivalente al bosque ombrófilo mesotérmico) es la mayor. Este valor debe ser tomado con precaución ya que el número de replicaciones (5 parcelas) en este bioclima es reducido, lo mismo ocurre con el Bioclima húmedo premontano (5 parcelas) (equivalente al bosque ombrófilo submesotérmico).
El valor promedio de biomasa del bioclima húmedo tropical (BHT) en todas las zonas geográficas de la cuenca del Caura es de 237 49 t/ha. Se puede apreciar que la menor biomasa promedio se presentan en las zona geográficas 6 y 14, la menor biomasa en 14 se atribuye a la presencia de suelos más pobres derivados de rocas de la Provincia Roraima (Rincón y Estanga 1996). El rango obtenido se corresponde con el intervalo estimado de densidad de biomasa aérea para los bosques tropicales de 200-500 t/ha (Cannel 1982, Tabla 1); también muestra magnitudes comparables a las obtenidas por el inventario RADAMBrasil con un promedio de 227 t/ha (166-332, ver Tabla 1) (Brown & Lugo 1992). Es de resaltar que los proyectos de inventario de CVG-TECMIN y RADAM-Brasil aplicaron metodologías y enfoques similares para sus inventarios de extensas regiones, asimismo los valores de biomasa del RADAM-Brasil se estimaron con la misma metodología aplicada en este trabajo (Brown & Lugo 1992), por lo cual se presume que su comparación sea bastante confiable.
Biomasa aérea y tipos de bosque
Al analizar la distribución de biomasa aérea por tipos de bosque (Tabla 6), el promedio de los bosques medios densos (BMD) en el bioclima húmedo tropical es 234 57,9 t/ha, valor que como era de esperar superó al mostrado por el bosque bajo denso (BBD) de 212 20,5 t/ha. En cuanto a los extremos del conjunto de los diferentes tipos de bosque se puede apreciar que la menor biomasa del BBD (197 31 t/ha) y una de las menores del BMD (205 26 t/ha) se presentan en la zona geográfica 14, región donde se presentan suelos desarrollados a partir de rocas de la provincia geológica de Roraima (Rincón y Estanga 1996, CVGTecmin 1995), lo cuales se presumen sean los de menor fertilidad de la cuenca.
El rango de valores de densidad de biomasa aérea obtenidos para los distintos tipos de bosques de la cuenca está dentro del amplio rango de 69- 550 t/ha para bosques del Escudo de Guayana (ver Tabla 1).
Al comparar los resultados de las estimaciones de densidad de biomasa aérea según los enfoques a pequeña y gran escala (Tabla 2, Tabla 5 & Tabla 6), se aprecia (tal como afirman Brown & Lugo 1992) una diferencia notable entre sus resultados tanto a nivel de bioclima como a nivel de tipos de bosques, siendo menores los valores estimados a gran escala. Particular interés despierta el hecho de que los bosques con palmas constituyen la única excepción, donde el valor estimado a gran escala es mayor que el estimado a pequeña escala; esta discrepancia se atribuye a dos factores, el primero alude al error que se comete cuando se aplican relaciones alométricas arbóreas a una forma de vida no arbórea como es el caso de las palmas y el segundo a la ocurrencia en la Cuenca del Caura de al menos dos grandes tipos de bosques con palmas, un tipo inundable y otro tipo que crece sobre tierra firme.
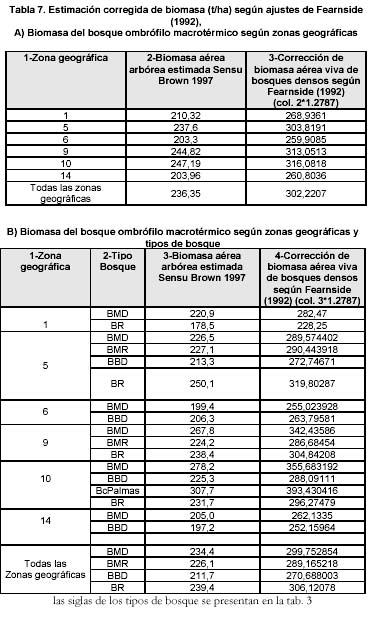
Ajustes de la anterior estimación de Biomasa aérea sensu Fearnside
Otra manera de abordar la estimación de otros componentes de la biomasa, diferentes a la biomasa aérea arbórea, consiste en aplicar los ajustes propuestos por Fearnside (1992, Tabla 4) para corregir las supuestas subestimaciones (ver Tabla 7) del modelo de Brown & Lugo (1992). Al realizar dichos ajustes es posible observar (ver Tabla 7) un incremento de los valores estimados de biomasa aérea anteriormente presentados. En el caso del valor promedio de biomasa aérea estimado inicialmente para los bosques en el bioclima húmedo tropical de 237 t/ha se transforma en 302 t/ha. En el caso del bosque medio denso en el bioclima húmedo tropical la media de 234 t/ha se transforma en 300 t/ha y para los bosques bajos densos en el bioclima húmedo tropical valor 212 t/ha se convierte en 271 t/ha y los bosques ribereños de 239 t/ha pasa a 306 t/ha, valores los cuales se acercan notoriamente a los resultados obtenidos en el estudio a pequeña escala de Chanaro (ver Tabla
2) para los tipos de bosque equivalentes.
Consideraciones finales
El conjunto de bosques aquí estudiados muestra una gran heterogeneidad, y abarca gran parte de la variación natural de los bosques del Escudo Guayanés. Este grupo de bosques se mantiene en rangos de biomasa similares a los estimados para otros bosques del Escudo de Guayana y de la Amazonia. Este hecho alude no sólo a las condiciones ambientales similares, sino también a su estrecha relación biogeográfica (Huber 1995).
Son muchos los factores que afectan la estimación de biomasa, entre ellos se pueden señalar la falta de ecuaciones adaptadas al área de estudio, las alometrías atípicas de formas de vida no arbóreas, sesgo en el muestreo, falta de replicación, la subestimación o subestimación de árboles muy grandes (Clark et al. en prensa). En este trabajo se presentan al menos los tres primeros factores, sobre todo por el hecho de haberse hecho las estimaciones haciendo uso de información preexistente resultante de estudios e inventarios, cuyos objetivos no fueron la estimación de biomasa. No obstante, el procedimiento aquí aplicado se considera válido como una primera aproximación de bajo presupuesto para estimar biomasa en bosques extensos. De cualquier manera, es evidente la necesidad de mejorar los niveles de precisión en la estimación de biomasa aérea en bosques del Caura Para lograr tal cometido un necesario próximo paso es el desarrollo de ecuaciones alométricas adaptadas a las condiciones y características de los bosques regionales a partir de información de biomasa colectada directamente en el terreno.
Para estudios futuros, de acuerdo a los recursos disponibles se podría en cada estrato (definido por zona geográfica, bioclima y tipo de bosque) desarrollar en el terrreno ecuaciones de biomasa locales fin de incrementar la exactitud en la estimación de densidad de biomasa aérea.
 |
|---|
La falta de exactitud en la estimación de la biomasa no solamente es una limitación en el componente arbóreo sino también en los otros componentes y en los flujos de biomasa. En el Caura, por ejemplo, no existen datos sobre la producción de necromasa. Asimismo aún son numerosas las especies en el Caura, cuya densidad de madera permanece desconocida. Por otra parte, la biomasa subterránea y la materia orgánica del suelo permanecen aún entre los aspectos más desconocidos.
Agradecimientos
Deseo agradecer a Astur Demartino por la colaboración en el acopio de información, a Hans ter Steege por la valiosa información suministrada, a Judith Rosales por su colaboración en el por su ayuda en la recolección de información. Este estudio pudo ser realizado gracias al financiamiento de BITOR-PDVSA y el auspicio de la Sociedad Conservacionista Audubon
de Venezuela.
 |
|---|
Bibliografía
Aymard, G., Elcoro, S., Marín, E., Chaviel, A. (1997). Caracterización estructural y florística en bosques de tierra Firme de un sector del bajo río Caura, Estado Bolivar, Venezuela. Estudios Especiales T.2. O. Huber, J. Rosales (eds.). Scientia Guianae 7:143- 170
Bongers, F, Engelen, D. & Klinge, H. (1985). Phytomass structure of natural plant communities on spodosols in southern Venezuela. the Bana woodland. Vegetatio 63: 13-34.
Briceño, J. (1995). Análisis fitosociológico de los bosques ribereños del río Caura en el sector Ceiato – Entrerios, Distrito Aripao del Estado Bolívar. Informe de Pasantía, Universidad de Los Andes, Facultad de Ciencias Forestales, Escuela de Ingeniería Forestal (CVG-EDELCA). Mérida. 90 pp.
Briceño, E., L. Balbas, J. Blanco. (1997). Bosques ribereños del bajo Río Caura. vegetación, suelos y fauna. En. Ecología de la Cuenca del Río Caura, Venezuela. estudios especiales, O.Huber & . Rosales (eds.). Scientia Guaianae 7: 259-289.
Brown, S. (1997). Estimating biomass and biomass change of tropical forests. a primer. FAO Forestry Paper 134. A Forest Resources Assesment publication. Rome 55 pp.
Brown, S. and A. Lugo. (1992). Aboveground biomass estimates for tropical moist forests of the Brazilian Amazon.. Interciencia vol. 17 1: 8-18.
Brown, S., Gillespie, A.J.R. & Lugo, A.E. (1989). Biomass estimation methods for tropical forests with applications to forest inventory data. Forest Science 35: 881-902.
Brown, S., Gillespie, Lugo, A. (1992). Tropical Forest Biomass Estimation From Truncated Stand Tables. Forest Ecology and Management. nº48 , Elsevier Science Publishers B.V. Amsterdam.
Brown, S. & A. Lugo. (1984). Biomass of tropical forests. a new estimate based on forest volumes. Science vol. 223: 1290-1293.
Bruenig, E.F. (1996). Conservation and Management of Tropical Rainforests, An integrated approach to sustainability. CAB International. 360 pp. Wallingford UK
Cannel, M. (1982). World Forest Biomass and Primary Production Data. Compiled by. M.G.R., Canell. Academic Press. London.
Castellanos, H. (1997). Ecología del comportamiento alimentario del marimona (Ateleus belzebuth Geoffrey, 1806) en el río Tawadu, Reserva Forestal El Cauara. En. Ecología de la Cuenca del Río Caura, Venezuela. estudios especiales, O.Huber & J. Rosales (eds.). Scientia Guaianae 7: 309-341.
Chudnoff, M. (1980). Tropical timbers of the world (Us. Dpto, of Agric., for service, for products Laboratory. Madison Wisconsin.
Clark, D.A. , S. Brown, D. Kckilighter, J. Chambers, J. Thomlinson & J. Ni & E. Holland (en prensa). Measuring Net Primary Production in forests. Concepts and field methods. Ecological Applications.
CVG-TECMIN C.A. (1995). Informe de avance NA-20-2. Clima, Geología, Geomorfología, Suelos y Vegetación. Tomo I. Proyecto inventario de los Recursos Naturales de la Región Guayana. Ciudad Bolívar. 634 pp.
CVG-TECMIN C.A. (1994). Informe de avance NB-20-10. Clima, Geología, Geomorfología, Suelo y Vegetación. Tomo I. Proyecto inventario de los Recursos Naturales de la Región Guayana. Ciudad Bolívar. 620 pp. Mapas escala 1:250.000.
CVG-TECMIN C.A (1994). Informe de avance NB-20-1. Clima, Geología, Geomorfología, Sulos y Vegetación. Tomo I. Proyecto inventario de los Recursos Naturales de la Región Guayana. Ciudad Bolívar. 557 pp.
CVG-TECMIN C.A. (1994). Informe de avance NB-20-9. Clima, Geomorfología, Geología, Suelos y Vegetación. Tomo I.- Proyecto inventario de los Recursos Naturales de la Región Guayana. Ciudad Bolívar. 512 pp.
CVG-TECMIN C.A. (1994). Informe de avance NB-20-13. Clima, Geología, Geomorfología, Suelos y Vegetación. Tomo I. Proyecto inventario de los Recursos Naturales de la Región Guayana. Ciudad Bolívar. 515 pp.
CVG-TECMIN C.A. (1994). Informe de avance NB-20-5. Clima, Geología, Geomorfología, Suelos y Vegetación. Tomo I. Proyecto inventario de los Recursos Naturales de la Región Guayana. Ciudad Bolívar. 606 pp.
CVG-TECMIN C.A. (1994). Informe de avance NB-20-14. Clima, Geología, Geomorfología, Suelos y Vegetación. Tomo I. Proyecto inventario de los Recursos Naturales de la Región Guayana. Ciudad Bolívar. 501 pp.
CVG-TECMIN C.A. (1991). Informe de avance NB-20-6. Clima, Geología, Geomorfología, Suelos y Vegetación. Tomos I y II. Proyecto inventario de los Recursos Naturales de la Región Guayana. Ciudad Bolívar. 1056 pp.
Dezzeo, N. & E. Briceño. (1997). La vegetación en la cuenca del río Chanaro. Medio río Caura. En. Ecología de la Cuenca del Río Caura, Venezuela. estudios especiales, O. Huber & . Rosales (eds.). Scientia Guaianae 7: 365-385.
Dezzeo, N., Briceño E., Hernández, L., Sanoja, E., Román, A., Rivas, Y. (1993). Informe de avance sobre las características de la vegetación boscosa en algunos sectores del área de inundación del proyecto “Trasvase Caura – Paragua”. Dpto. De Estudios Básicos – EDELCA, Dpto. De Conservación de Cuencas – EDELCA, Dpto. de Botánica-UNEG. Puerto Ordaz 15 pp.
Díaz, J & Pernía, H. (1995). Caracterización de la vegetación de dos sectores de la cuenca alta del río Caura (entre 5° y 5° 25’ latitud Norte y 63° 45’ y 64° 22’30” longitud oeste). Informe de pasantía. Universidad de los Andes, Facultad de Ciencias Forestales. Escuela de Ingeniería Forestal. Mérida. 117 pp. + tablas.
Fearnside, P. (1997). Wood density for estimating forest biomass in Brazilian Amazonia. Forest Ecology and Management. 90 (1997) 59-87.
Fearnside, P. M. (1992). Forest biomass in Brazilian Amazonia. Comments on the estimate by Brown and Lugo. Interciencia vol. 17 1: 19- 27.
Fuentes, J. & J. Madero (1996). Suelos. En. Ecología de la Cuenca del Río Caura, Venezuela I. Caracterizacòn General . J. Rosales & O. Huber (eds.) Scientia Guaianae, 6: 44-47. Caracas.
Gérard, J., Millar, R., B. Ter Welle (1996). Major timber trees of Guyana. Tropenbos series 15. The tropenbos foundation. Wageningen. The Netherlands 224 pp.
Hase, H., Fölster, H. (1982). Bioelement inventory of a tropical (semi) evergreen seasonal forest on eutrophic alluvial soils, Western Llanos, Venezuela. Acta Oecológica/Oecológica Plantarum vol. 3(17) N°4.
Holdridge, L. (1947). Determination of world plant formation fron simple cliamte data. Science 105 (2727): 367- 368.
Huber, O. (1996). Vegetación no boscosa. En. Ecología de la Cuenca del Río Caura, Venezuela I. Caracterización General . J. Rosales & O. Huber (eds.) Scientia Guaianae, 6: 7-75. Caracas.
Huber, O. (1995). Vegetation, chapter 3 in. Flora of the Venezuelan Guayana. Vol. I. Introduction. By P. Berry, B. Holst and K. Yatskievych (vol. eds.). J. Steyermark, P. Berry and B. Holst editores generales de la serie. 97-160.
Huber, O. & J. Rosales (eds.) (1997). Ecología de la Cuenca del Río Caura, Venezuela II. Estudios Especiales. Scientia Guaianae, N° 7. Caracas. 468 pp.
Huber, O., J. Rosales & P. Berry (1997). Estudios botánicos en las montañas altas de la cuenca del Rio Caura (Edo. Bolívar, Venezuela). En. Ecología de la Cuenca del Río Caura, Venezuela II. Estudios especiales. O. Huber & J. Rosales (eds.) Scientia Guaianae, N° 7: 441-468. Caracas.
Jordan, C.F. ed. (1989). An Amazonian rain forest. The structure and function of a nutrient stressed ecosystem and the impact of slashand- burn agriculture. MAB series 2. Parthenon Publishing Group.
JUNAC- PADT. REFORT (1981). Tablas de propiedades físicas y mecánicas de la madera de 20 especies de Venezuela. Junta de Acuerdo de Cartagena. JUNAC- PADT. REFORT 53 pp.
Klinge, H. & Herrera, R. (1983). Phytomass structure of natural plant communities on spodosols in southern Venezuela. the tall Amazon Caatinga forest. Vegetatio 53: 65-84.
Klinge, H., & Medina, E. (1979). Rio Negro caatingas and campinas, Amazonas states of Venezuela and Brazil. In. Specht, R.L. Heathlands and related shrublands. Ecosystems of the World 9A. Elsevier, Amsterdam.
Klinge, H., Rodrigues, W.A., Bruenig, E. & Fittkau, E.J. (1975). Biomass and structure in a central Amazonian rain forest. In. Golley, G.B. & Medina, E. (eds.) Tropical ecological systems. Springer, New York.
Knab-Vispo, C. (1998). A rain forest in the Caura reserve (Venezuela) and its use by the indigenous Ye’Kuana people. Thesis, Doctor of Philosophy (Land Resources, University of Wisconsin – Madison. 202 pp.
Marin, E & Chaviel (1996). Bosques de tierra firme. En. Ecología de la Cuenca del Río Caura, Venezuela I. Caracterizacòn General . J. Rosales & O. Huber (eds.) Scientia Guaianae, 6: 60-65
MARN. (2000). Indice de cubrimiento cartográfico 1935-2000. MARN, Servicio Autónomo de Geografía y Cartografía Nacional. Caracas. 16 p.
Miranda M., L. Hernández, J. Ochoa, E. Yerena & A. Blanco-Uribe. (1998) «All That Glitters is Not Gold. Balancing Conservation and Development in Venezuelas Frontier Forests». Word Resource Institute. Washington DC. 53 pp. Trad. al español.
Phillips, O., Y. Malhi, N. Higuchi, W. Laurance, P.V. Nuñez, R.M. Vásquez, S. Laurance, L.Ferreira, M. Stern, S. Brown & J. Grace (1998). Changes in the carbon balance of tropical forests . evidence of long term plots. Science vol. 282: 439-442.
Rincón, H. & Y. Estanga (1996). Geología. En. Ecología de la Cuenca del Río Caura, Venezuela I. Caracterizacòn General . J. Rosales & Huber (eds.) Scientia Guaianae, 6: 20- 24. Caracas.
Rosales, J. & O. Huber (eds.), (1996). Ecología de la Cuenca del Río Caura, Venezuela I. Caracterización general. Scientia Guaianae, N° 6. Caracas. 140 pp.
Rosales, J., C. Knab – Vispo y G. Rodríguez. (1997). Bosques ribereños del bajo Caura entre el Salto Pará y los Raudales de La Mura. Su clasificación e importancia en la cultura Yekuana. En. Ecología de la Cuenca del Río Caura, Venezuela II. Estudios Especiales. O. Huber & J. Rosales (eds.) Scientia Guaianae, 7: 171-213. Caracas.
Rosales, J. (2000). Re porte sobre los bosques ribereños del río Caura. Proyecto Bitor. 13 pp. Manuscrito inédito.
Rosales, J. (2000). An ecohydrological approach for riparian forest biodiversity conservation in large tropical rivers. PhD Thesis University of Birmingham. 208 pp.
Rosales, J, C. Vispo, N. Dezzeo, L. Blanco, C. Knab-Vispo, N. González, C. Bradley; D. Gilvear, G. Escalante, N. Chacón and G. Petts (en prensa). Riparian forests ecohidrology in the Orinoco River Basin. In. The Ecohidrology of South American Rivers and Wetlands (McClain et al. eds.). UNESCO IHP Ecohidrology. Ecohidrology Programme
Salas, L., P. Berry & I. Goldstein (1997). Composición y Estructura de una comunidad de árboles grandes en el valle del Rio Tabaro, Venezuela. Una muestra de 18,75 has. En. Ecología de la Cuenca del Río Caura, Venezuela II. Estudios Espéciales. O. Huber & J. Rosales (eds.) Scientia Guaianae, 7: 291-308. Caracas.
Por: Lionel Hernández
Fuente: Revista Forestal Iberoamerica